
私達は、植物らしさのひとつである環境因子による発生過程の柔軟な細胞増殖や分化制御の分子実体の解明や制御機構の解明を目指しています。分子遺伝学を主体としながら、遺伝子、分子、細胞、個体、生態系の各レベルの垣根を超えた研究を目指しています。研究科HPの紹介ページもご覧ください。[English page]
研究領域:植物の光環境応答、生長制御、有性生殖、性決定、性分化、陸上植物進化、比較ゲノム
成果紹介HPへのリンク
植物の永続的な成長を支える分子機構を解明~成長点の司令塔を担う転写因子が鍵~ [HP]
雌雄異株から雌雄同株への進化に伴う性染色体の運命とは―コケ植物の有性生殖システム転換における染色体再編成の解明― [HP]
コケ植物の卵細胞を生み出す遺伝子を発見―陸上植物の生殖細胞をつくる機構とその進化― [HP]
植物の生殖細胞をつくる2つの遺伝子―5億年前に誕生したヘテロ二量体― [HP]
コケで探る植物ホルモンジベレリンの多様性―苔類ジベレリン様化合物は遠赤色光応答を制御― [HP]
ゼニゴケを用いてオーキシンの働きを証明ーオーキシンなくして器官形成なしー [HP]
ゼニゴケは遺伝子のオモテとウラを使ってメスとオスを作り分けている~性差を生み出す巧妙な「裏ワザ」が明らかに~ [HP]
植物の生殖細胞をつくる鍵因子を発見 -花粉の精細胞をつくる仕組みは花の咲かないコケ植物に起源があった-
植物のストレス応答に重要な植物ホルモンであるジャスモン酸信号伝達系の原型
陸上植物の季節に依存して花を咲かせる仕組みの起源を紐解く [HP]
(研究費紹介ページ)
陸上植物の性分化:遺伝的頑健性と可塑性のメカニズム (基盤S)
苔類ゼニゴケをモデルとした分子遺伝学の展開
我々は、新たなモデル植物として、苔類ゼニゴケに注目しています。生活史の大半が半数体であること、陸上植物の起源を考える上で進化系統的に鍵となるなど興味深い材料です。我々の研究室では、アグロバクテリアによる高頻度形質転換とプラスチドの形質転換に成功しています。また、実験室環境で世代をまわすことも可能になり、雌雄異種の半数体であることを生かした遺伝学展開が可能と考えています。現在、ゲノム遺伝地図を作製しつつあり、ゲノム解読も米国JGIとの共同研究でスタートしました。制御因子に関しては、基本的には陸上する因子を保持するものの、その遺伝的冗長性の低さは際立っています。遺伝的な利点を生かすべく、ガンマ線照射やT-DNAタグによる突然変異体単離もはじめました。相同性組換えによる遺伝子ターゲティングをはじめ、実験のツールは急速に整備されつつあります。ゼニゴケのもつ単純さ、細胞可視化の容易さ、世代時間の短さといった特徴を生かした新しい分子遺伝学研究が始まりました!
特定領域研究【オルガネラ分化】のニュースレター7号(2008.1発行)に、実験生物としてのゼニゴケの紹介記事を書きました(読む)。ただし、ゼニゴケに注目した初期の記事なので内容的にはその後の進展があります。
また、日本植物生理学会の一般向けページ、みんなのひろばでも「モデル植物としてのゼニゴケ(読む)」というタイトルで解説をしています。
日本植物学会『植物科学の最前線(BSJ-Review)』 では、我々のゼニゴケを扱った和文総説集が収録公開されています。以下のタイトルで書いていますので、我々がどのようなことに興味をもって研究を進めているかの参考にしてください。
1 古くて新しいモデル植物としてのタイ類ゼニゴケの特徴 河内孝之・石崎公庸 BSJ-Review 3:58-70 (2012) (読む)
2 見えてきたゼニゴケゲノム 大和勝幸・河内孝之 BSJ-Review 3:71-83 (2012) (読む)
3 基部陸上植物の光応答戦略 -フィトクロムを介した光形態形成の分子機構- 石崎公庸・井上佳祐・河内孝之 BSJ-Review 4:37-44 (2013) (読む)
4 陸上植物の細胞分裂の光制御とその進化 西浜竜一・河内孝之 BSJ-Review 6A:51-62 (2015)(読む)
5 植物における日長による成長相転換制御のメカニズムとその進化 山岡尚平・河内孝之 BSJ-review7B:78-86 (2016)(読む)
更に興味をもたれた方は、直接我々にお問い合わせください。
ゼニゴケに関して研究室で進行中の研究は、次の通りです。
- 赤色光・遠赤色光受容体phyとシグナル伝達
- 光質と日長による有性生殖誘導機構
- ジベレリン様化合物による信号伝達の進化的変遷
- ゼニゴケジベレリン様化合物の分子実体と生合成経路の解明
- 半数体生物の性決定遺伝子による性決定の分子機構
- 有性生殖のマスター転写因子の発現制御機構 など
参考のため、これまでに行った研究も掲載しています。
これまでに行った研究
植物の光受容と信号伝達に関する研究
我々は、植物の主要な光受容体のひとつであるフィトクロムの発色団フィトクロモビリンに注目し、その分子構造と光受容能および個体の生理応答を解析している。シロイヌナズナの光形態形成変異体を材料に、フィトクロム発色団の生合成に関わる遺伝子群を単離同定した。HY1遺伝子はヘ ムからビリベルジンへの開環反応を触媒するヘムオキシゲナーゼを(1, 2)、HY2遺伝子はビリベルジンからフィトクロモビリンの還元反応を触媒するフィトクロモビリン合成酵素をコードしていた(3)。 両者は、フェレドキシン依存の還元酵素で葉緑体に局在した。
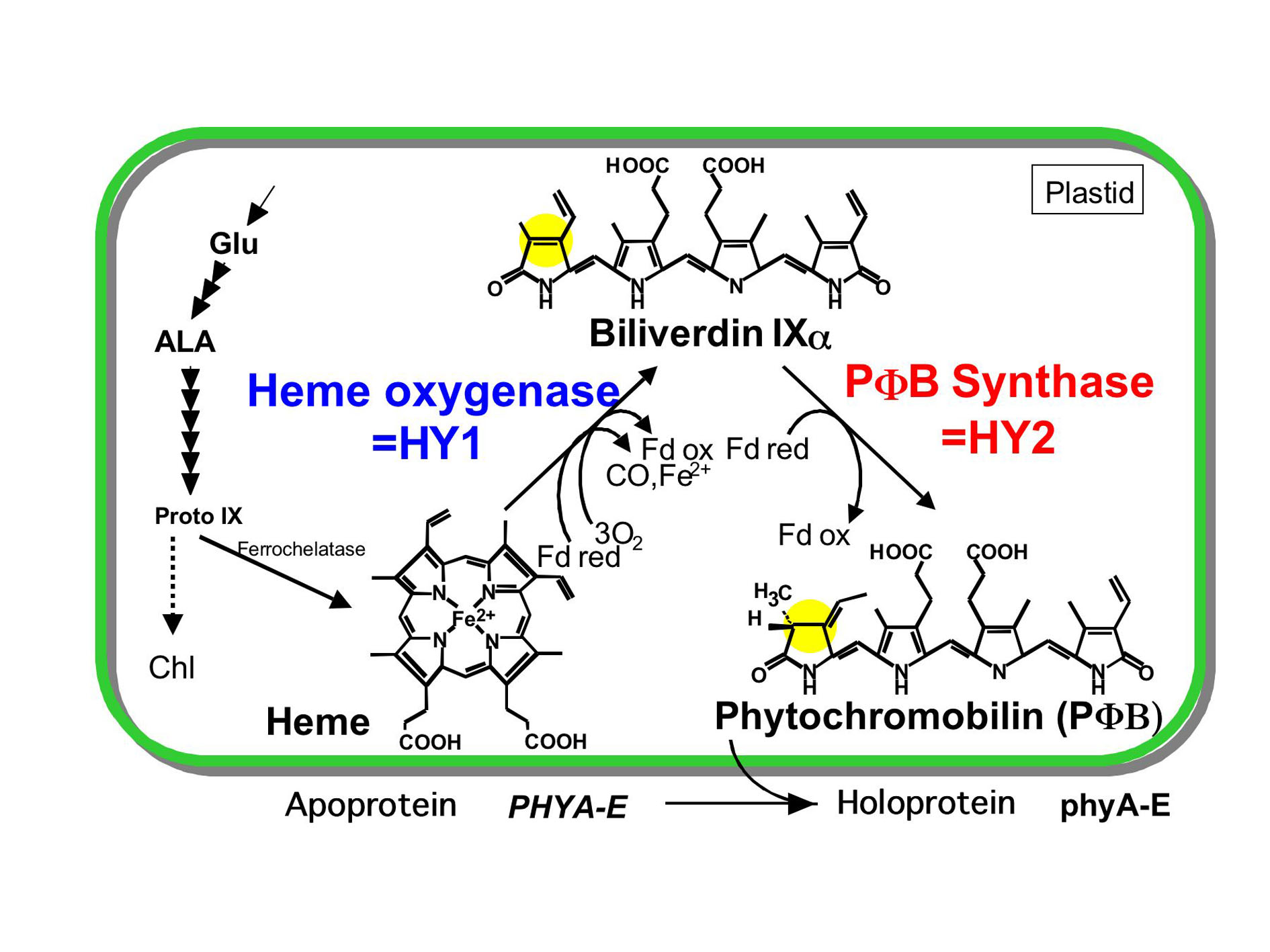
また、HY2に対する相同性に基づき、藍藻からフィコビリソームの成分であるフィコシアノビリン(PCB)やフィコエリスロビリ ンの合成に働くビリン還元酵素遺伝子を同定した(4)。 単離した遺伝子とhy2突然変異体を利用して、高等植物のフィトクロム発色団構造を藍藻フィトクロム様タンパク質の発色団であるPCBに改変する系を構築し、発色団と生理応答の機能的な相関を解析した。その結果、発色団構造は、遠赤色光の波長特異性などを規定するのに重要であることがわかった(5)。 これまでにトマト(6) とトウモロコシ(7) のフィトクロモビリン合成酵素欠損変異体を同定できたので、同様の手法で作物で発色団の改変することも現実的である。以上のように、これまでの研究では、 フィトクロムの光受容とシグナル伝達経路の解明への基礎的な知見を蓄積するとともに、単離した遺伝子を活用して光受容体構造を改変することにより光に対する応答特性が変化するという植物の分子育種への応用の可能性を示すとこができた。
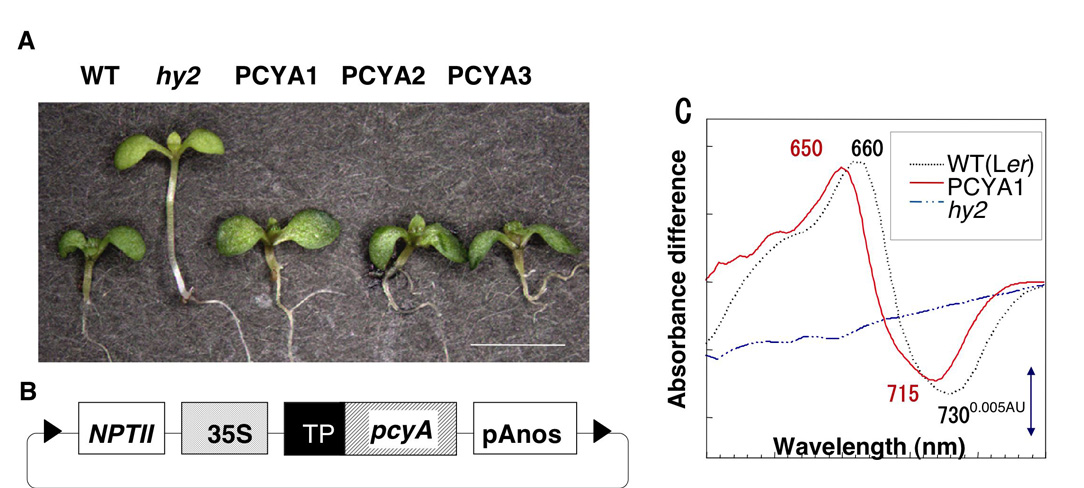
単離した遺伝子をもとにフィトクロム発色団生合成酵素とフィトクロムタンパク質を大腸菌で共発現させ、光受容能のあるフィトクロムを高発現する系を構築した。現在、世界中でシロイヌナズナ突然変異体を用いて光信号伝達の遺伝的経路の解析が盛んに行われているが、その分子機構の全貌の解明にはほど遠い。今後は、我々の構築したフィトクロム発現系を有効に活用し、フィトクロム信号伝達の初期反応を解明するため、相互作用因子の解析を進める計画である。分子遺伝学的手法に加えて、プロテオミクスを積極的に取り入れたいと考えている。また、単純な光情報伝達経路を解析することが重要という考えから最古の陸上植物である苔類の光応答とフィトクロムに関する研究も開始した。我々の研究では、光受容という信号伝達の入力から遺伝子発現制御に至る経路の分子的 実体を解明し、生物の光スイッチとしての高度利用に供することが究極的な目標である。
植物の花成制御に関与するMADSボックス転写因子に関する研究
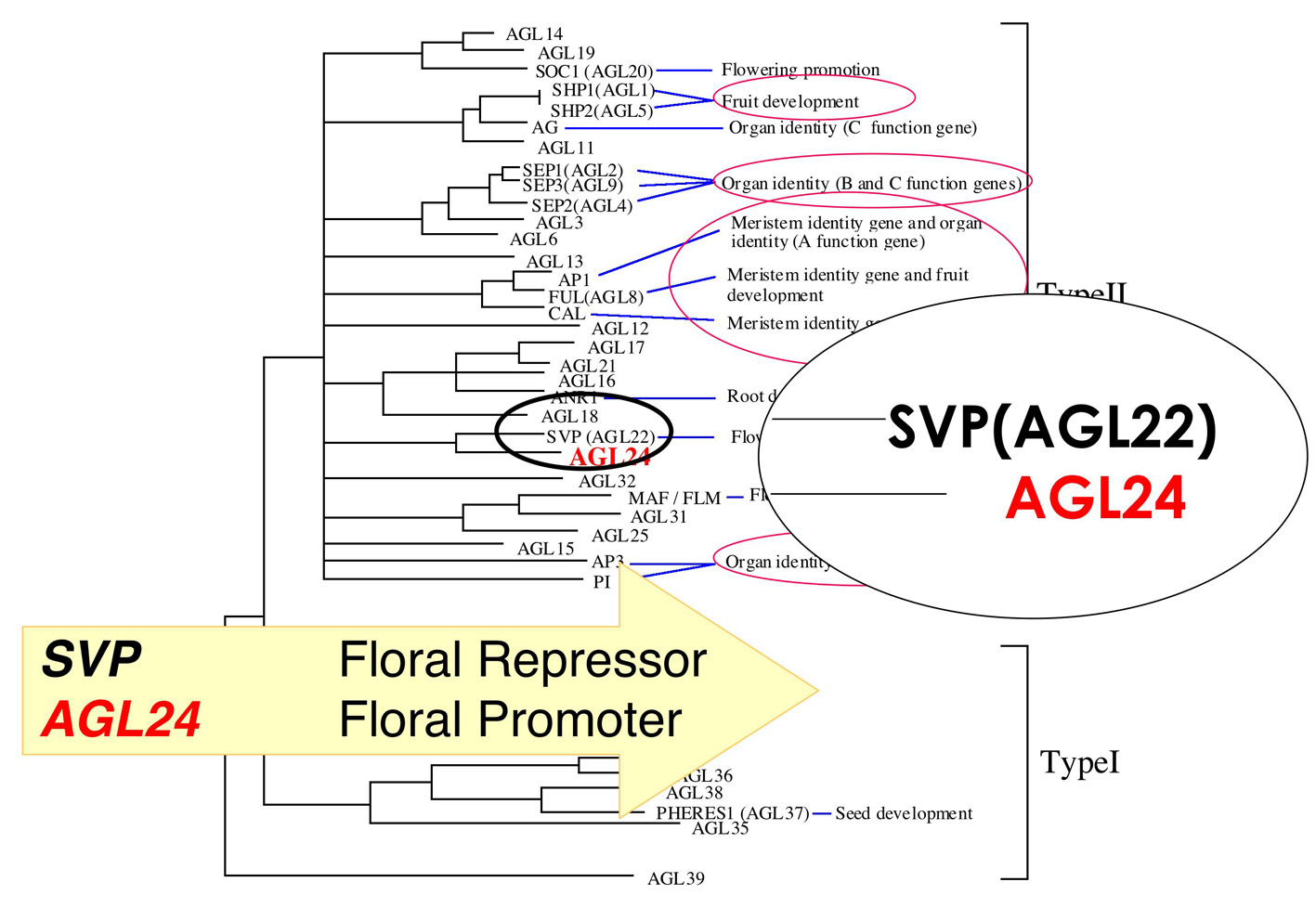
植物は頂端分裂組織の維持と分化により、胚発生以降も継続的に発生を繰り返す。植物の発生の特徴である分化全能性と無限生長は茎頂分裂組織の機能による。また、遺伝的プログラムと環境要因に依存する生長相転換により発生する器官が変化する。我々はシロイヌナズナを用いて頂端分裂組織特異的に発現する受容体キナーゼの標的タンパク質としてMADSボックス転写因子AGL24を同定した(8)。 AGL24は栄養成長から生殖成長への移行(花成)の促進因子である。興味深いことに、配列的に最も近縁なSVPは花成の抑制因子である。花成の誘導は、 環境要因などが葉で感知されることが知られているが、花器官の発生は分裂組織を介した出来事である。AGL24は茎頂で発現するタンパク質であり、花成誘導後のシグナルが統合されて茎頂で発生する器官を規定する役割が想定される。現在、AGL24とSVPの形成する複合体分子の解析や形質転換手法による分子遺伝学的な表現型解析によって、花成経路の下流における遺伝子相互の遺伝学的な関係と、分子としての機能発現を規定する機能的な分子構造を明らかにすることを目指している。
葉緑体ゲノムの複製と維持に関する研究
葉緑体は独自のゲノムをもち、100ほどの重要な遺伝子をコードしている。葉緑体機能を発現するには核の支配、すなわち核にコードされる遺伝子の機能が必須であることが強調されがちであるが、炭酸固定のキー酵素RubisCOのLサブユニットをはじめ、葉緑体にコードされる遺伝子発現は植物の生存のために極めて重要である。葉緑体(色素体)ゲノムは主として分裂組織や器官原基で複製し、そのコピー数は組織や光条件により異なる。葉緑体の発達した細胞では、葉緑体の数と葉緑体ゲノムのコピー数の増加により、10,000コピーもの葉緑体ゲノムが存在する。葉緑体DNA上にその制御の標的が存在すると考えられたが、オルガネラゲノムの配列からは複製制御に関わる配列は予想されなかった。我々は、葉緑体ゲノムの複製制御メカニズムに興味を持ち、ゼニゴケを材料に高効率の葉緑体形質転換手法を開発した。